

Managing Sandplain Grasslands in a Brave New World
The Sandplain Grassland Network compiled and reviewed lessons learned from the past several decades of sandplain grassland science and management. While the lessons from previous experiences are important, future management to maintain sandplain grasslands as regional biodiversity hotspots will likely involve new challenges. This is because sandplain grasslands within northeastern U.S. coastal landscapes will increasingly be influenced by climate change, spread of new or different non-native and invasive species, changing patterns of land use, herbivore populations, and attitudes toward land conservation and land management. Recognizing these challenges—and facing up to what we do and do not know about how this “brave new world” will influence sandplain grasslands—will help to guide future management and research, and improve the success of future sandplain grassland management.
Climate Change
Although climate change already affects the northeast U.S., in the upcoming decades these impacts will be severe and will increasingly influence sandplain grassland biota. The climate of the region has warmed 1.3 oC (2.4 oF) since 1895, and the annual mean temperature exceeded the 20th-century average every year since 1993 (Bradley et al. 2018). Between now and about 2065, the average summer and winter temperatures in Massachusetts will likely increase more than 3.3 oC (6 oF) relative to pre-industrial levels. The coldest winters of the future will be like the warmest winters of recent years and the coolest future summers will be like our hottest summers of today. Summer in the northeast U.S. by the end of the century will feel like a present-day typical summer in South Carolina. Annual rain and snow has already increased about 200 mm (7.8 in) since 1900 and the amount of precipitation falling during intense multi-day events increased by 71% between 1958 and 2012—a greater increase in intense precipitation than anywhere else in the U.S. The future will bring more winter rain and less winter snow. The future of summer rainfall is less certain but summer rain will likely become more variable so that more intense summer droughts may occur even while total yearly precipitation rises (Horton et al. 2014). We currently know relatively little about how these climate changes will affect most of the important plant and animal species that sandplain grasslands support.
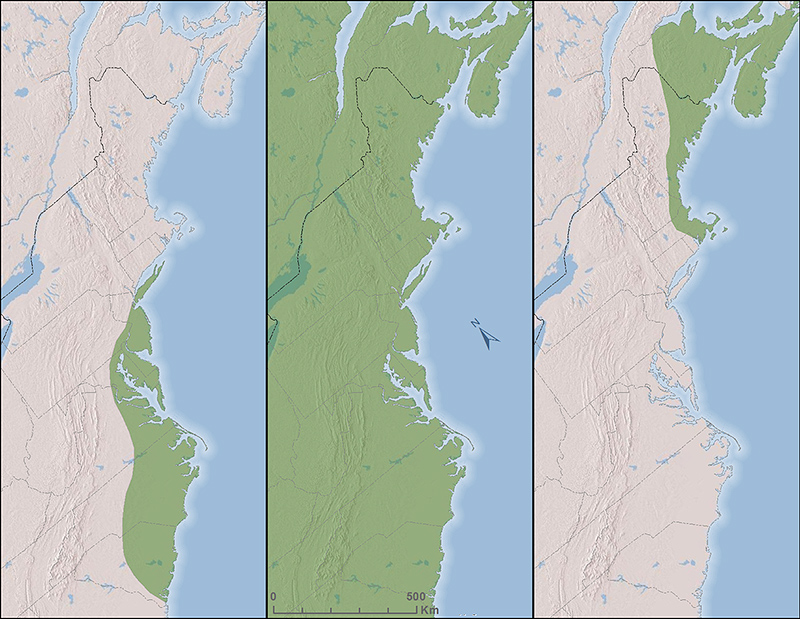
Figure 1. Hypothetical examples of sandplain grassland species with southerly (left), widespread (middle) or northerly (right) current ranges.
Warmer temperatures alone will likely shift the distributions of many sandplain grassland species over time. This may make it harder or easier to manage for different species depending on the north-south position of their current ranges (Fig. 1). Species near the current northern edge of their range may increase and may become easier to manage for in the future. For example, Blue Grosbeak (Passerina caerulea) is a grassland and shrubland bird species with a current southerly North American distribution. It is increasing in the northeast and was first successfully bred in Massachusettsetts in 2017 (Neill 2017). In contrast, grassland species now at the southerly edge of their range may become harder to manage for and face greater risk of regional extirpation. Broom crowberry (Cormea conradii) is an Ericaceous shrub that has a restricted current coastal range from New Jersey north to Newfoundland. Management of broom crowberry could become much more challenging in the future especially within the southern part of its range. How climate change influences the management of other important common sandplain grassland species that have very broad ranges may be less pronounced. The ranges of foundational sandplain grassland species such as little bluestem grass (Schizachyrium scoparium) or Pennsylvania sedge (Carex pensylvanica) extend well to the north and south of the northeast U.S., so their future ranges will almost certainly continue to include this region under the projected near-term future climate. These potential range shifts will almost certainly not be restricted to sandplain grassland species of conservation interest, but will potentially occur also for woody or invasive species that create persistent challenges for sandplain grassland managers. A comprehensive evaluation of current and potential future ranges of target and problem species, which could provide information on the potential importance of these shifts, has not been conducted.
Future climate changes that bring more variable precipitation and more severe occasional summer droughts could affect the ecological dynamics and management of sandplain grasslands in several important ways. Because sandplain grasslands occur on well-drained soils, effects of flooding caused by more extreme rainfall will likely be minor. However, drought might decrease seed set and increase the years in which many sandplain grassland plants either produce fewer seeds or lower quality seed. This could influence approaches to restoration actions that depend on seed collections from existing grassland-associated plants. If the frequency and severity of droughts and dry summers increases, it could also reduce seed germination or the survival of seedlings of plants planted into restoration projects. More frequent or severe droughts will likely make predictable management by prescribed fire more difficult by creating more “red flag warning” days, and fewer days when fire can be used during the growing season, when fire is most effective for controlling growth of woody plants in sandplain grasslands. In contrast, increased frequency of dry summers might increase the effectiveness of fire during the growing season, at the times when it can be used, because the combined stress of fire and drought could reduce the survival and re-sprouting of the remaining root stocks of woody plants.
Shifts toward earlier flowering because of warming temperatures are well documented in the northeast U.S. (Miller-Rushing and Primack 2008). These changes are a concern because they could disconnect the time of flowering from the time in which pollinators are active, thus disrupting both plant and pollinator reproduction and their long-term viability (Scaven and Rafferty 2013). These changes have not been investigated specifically in sandplain grasslands. Warming temperatures could also change conditions in locations in the coastal sandplain grassland region where microtopographic features influence the occurrence and timing of low temperatures, and the relative balance between trees and a shrubbier community that contains a larger grassy component. On sandy and drought-prone soils, topographic lows can result in “frost pockets” where nighttime below-freezing temperatures occur into early summer (Aizen and Patterson 1995). Because these late frosts typically occur before scrub oak (Quercus illicifolia) but after tree oak (Q. alba, Q. velutina) leaf out, they can cause greater tree oak mortality and maintain scrub oak barrens (Motzkin et al. 2002). Warmer springs or cloudier conditions that prevent radiative nighttime cooling would potentially eliminate this phenomenon and increase the presence of woodland at the expense of scrub oak shrubland that frequently harbors at least some plant species more typical of sandplain grasslands.
Rising sea levels already influence the conservation management of sandplain grasslands and the effects of rising sea levels will increase in the future. Many excellent examples of native species-rich sandplain grasslands occur directly behind the open Atlantic Ocean coastal dunes or adjacent to coastal salt and freshwater ponds, particularly on Long Island, the mainland coast of Massachusetts, Nantucket, and Martha’s Vineyard. In these locations, frequent disturbance by wind and salt spray helps to limit tree growth and maintain grass cover (Griffiths et al. 2006). This aids management because less aggressive efforts will be needed to maintain grass and prevent tree regrowth in strictly coastal sites compared with sites farther inland. Sea level rise already threatens these coastal sandplain grasslands that lie very near the coast. Sea level in the northeast has risen approximately 20 cm since 1900 and the rate of sea level rise is increasing. Recent studies indicate that Massachusetts will likely see 100 cm of additional sea level rise by 2100 and nuisance flooding of low-lying areas along the U.S. east coast is increasing (Sweet et al. 2018). This means that some of best current examples of sandplain coastal grasslands will likely experience more extreme storm surges and the accompanying effects of salt spray, temporary flooding, and wind damage during this period.
Imminent and accelerating sea level rise means that maintaining the most coastal sandplain grasslands in the northeastern U.S. will require the ability to: (1) allow current coastal sandplain grasslands to gradually migrate inland, or (2) create sandplain grasslands at locations farther inland in places that are currently other habitats. Both strategies will likely be necessary to sustain even the current area of high quality grasslands and the implementation of these strategies can benefit from lessons learned in this review. Although relatively few locations exist in the populated northeast U.S. coastal region where corridors of undeveloped lands exist in contiguous swaths that connect large, inland protected areas directly to the coast, some of the areas where this landscape connectivity exists are already important locations of sandplain grasslands. Large central portions of the island of Nantucket that include the Sanford Farm/Ram Pasture, Head of the Plains, Trots Hills, and Smooth Hummocks properties, contain excellent examples of coastal sandplain grasslands in locations that are contiguous with protected lands farther inland. There are other examples of these landscapes at the Katama Plains and the Long Point Wildlife Refuge on Martha’s Vineyard, and surrounding Allens Pond in southeastern Massachusetts. Even in these locations, the loss of sandplain grasslands at the coast will not necessarily be compensated for by expansion of sandplain grasslands inland into what are currently more shrub- or tree-dominated areas. A high long-term management priority for these still-contiguous properties to adapt to sea level rise might be to aggressively enlarge patches of existing sandplain grassland near the coast into these more wooded areas farther inland. In some instances, these contiguous inland areas are themselves highly-valued, terrestrial habitats such as scrub oak woodlands or pitch pine barrens that are conservation priorities. This will require careful assessment of tradeoffs by land managers. In many cases, more landscape-scale plans for maintaining or creating sandplain grasslands farther inland can draw on the experiences gained from managing woody regrowth in existing grasslands or converting existing shrublands or woodlands to grassland.
There is a much larger number of, typically smaller, examples of existing, highly coastal sandplain grassland patches, for which such protected migration pathways on protected lands do not exist. To compensate for the future loss of these areas to rising sea levels, managers should consider opportunities to create new sandplain grasslands on existing woodlands or existing anthropogenic grasslands in more inland locations.
Species Invasions
Many areas of current extent of sandplain grasslands were used for agriculture for some time between European settlement and the early- to mid-20th century (Motzkin and Foster 2002). Most sandplain grasslands have a component of their flora composed of non-native species because of this land use legacy. Compared with sites that were never tilled, grasslands that were formerly tilled generally have in their floras a number of non-native, widespread, weedy plants (Von Holle and Motzkin 2007).
Today, sandplain grasslands face increasing pressures from invasion by non-native and invasive species for several reasons. First, sandplain grasslands in the northeast U.S. now exist within a landscape, highly fragmented by residential development (Reinmann and Hutyra 2017) in which the proximity of grasslands to residential lands has greatly increased. The floras of residential yards in the northeast and elsewhere in the U.S. contain high numbers and proportions of non-native species and species that can easily colonize surrounding lands. Many of the invasive species that currently cause management concerns in sandplain grasslands, such as oriental bittersweet (Celastus orbiculatus), bush honeysuckles (Lonicera spp.), autumn olive (Elaeagnus umbellata) and multiflora rose (Rosa multiflora), have been present to various extents in some sandplain grasslands for many decades. However, persistent re-invasion of these species is more likely today that it was in the past because of increased dispersal of seeds that originate in the edge-rich residential landscapes in which sandplain grasslands are now embedded.
Second, the abundances of some invasive species that are relative newcomers in sandplain grasslands appear to be increasing rapidly in the northeast U.S. Three examples are Amur peppervine (Ampelopsis glandulosa), spotted knapweed (Centaurea stoebe), and black swallow-wort (Cynanchum louiseae). Reasons for the recent increase are not entirely clear, but it appears to be occurring in New York (including Long Island) and Massachusetts. Warmer temperatures and especially warmer winters may be factors, as well as the increase of carbon dioxide concentrations in the global atmosphere. Most vines, including poison ivy (Toxicodendron radicans), increase their growth rates faster than other plants in the presence of elevated carbon dioxide (Mohan et al. 2006).
Many sandplain grasslands also have a number of native species of vines and shrubs that persisted through periods of agricultural use, but then increased when lands were released from grazing or tilling. Some species likely persisted because they were protected from grazing by thorns (such as catbrier, Smilax rotundifolia; or brambles, Rubus spp.) or because they were unpalatable (such as black huckleberry, Gaylussacia baccata). Sandplain grasslands that were released from grazing relatively recently are likely undergoing a current rapid expansion of these species because spread occurs largely by clonal reproduction, and accelerates after an initial, relatively slow increase in the ratio of edges to open areas. The spread of these species into remaining grasslands on Naushon Island is an example of this recent rapid expansion of native shrub species into grasslands (Champlin 2016).
In unusual cases, the introduction or spread of an invasive species may create new opportunities for creating sandplain grasslands or shrublands in new places. The spread of the southern pine beetle (Dendroctonus frontalis) across the Pinelands in 2001 and onto Long Island in 2014 threatens the region’s pitch pine (Pinus rigida) forests because of the beetle’s effectiveness in killing trees usually within several months of infestation (Dodds et al. 2018). This range expansion will almost certainly continue in the northeast U.S. in coming years. The inevitable loss of pitch pine forests results in areas that could be considered for sandplain grassland creation and expansion.
Trophic Interactions
The proximity of sandplain grasslands to fragmented residential landscapes has consequences for many sandplain grassland-associated animal species. High densities of medium sized predators like skunks and raccoons occur in residential landscapes (DeStafano and DeGraf 2003) and are associated with sources of food, availability of denning sites in residential landscapes, and the loss of apex predators. Some of these predators, like crows, are highly mobile and can key in on specific habitats at certain times of year. The abundance of generalist predators in adjacent residential lands can influence sandplain grassland birds and reptiles, even when these predators do not specifically target grassland species of high conservation concern (Vickery et al. 1992). Domestic cats are likely the number one anthropogenic cause of wildlife mortality in the U.S. (Loss et al. 2013) and grassland birds nesting on or close to the ground are particularly vulnerable to predation (Isaksson et al. 2007). Because many sandplain grasslands are popular places for people in increasingly populated surrounding areas to walk, disturbances by dogs, particularly during the nesting season, are an increasing concern. Roads increase mortality of reptiles (Foreman 2002) and a higher density of roads near sandplain grasslands today compared with in the past almost certainly threatens box turtles (Terrapene carolina), black racers (Coluber constrictor), and eastern hognose snakes (Heterodon platirhinos) that use sandplain grasslands. These trophic interactions created by adjacent residential landscapes create new and persistent challenges for sandplain grassland management. Predator control is typically publicly unacceptable and highly controversial. Management of vegetation both in and around grasslands to promote target species, coupled with education targeted at surrounding residential landowners (Lutter et al. 2018), may be the best alternative approach to managing higher predation rates.
The populations of herbivorous white-tailed deer (Odocoileus virginianus) have also increased throughout the northeastern U.S. sandplain grassland region. Deer populations on Long Island, in Massachusetts, and other places are now above estimated carrying capacity and various approaches to reducing deer numbers are being considered. Deer herbivory can threaten sandplain grassland plants that are conservation targets. Some species such as lion’s foot (Nabalus serpentarius), are highly affected by deer browse and occur in numbers only in places like the Nantucket Airport that are fenced and where deer are controlled. Deer and rabbits also graze heavily on New England blazing star (Liatris scariosa var. novae-angliae) and damage to plants is greatest when grazing occurs during the middle of the growing season.
More of the current debate about high deer populations in the coastal sandplain region focuses on the role that deer play as hosts to the ticks that transmit Lyme disease, erlichiosis, anaplasmosis, babesiosis, rocky Mountain spotted fever, and other vector-borne diseases. Insect-borne diseases reported to the Center for Disease Control in the U.S. Mosquito and tick-borne diseases increased three-fold between 2004 and 2016 and are spreading rapidly in the northeastern U.S. While the increase in Lyme and other diseases has been attributed to an increase in deer populations (Wilson et al. 1988), this link is not straightforward above relatively low deer densities and may be more closely associated with the reductions in the numbers of predators that prey on the small mammals that are primarily responsible for transmitting the bacteria (Borrelia burgdorferi) that cause Lyme disease (Levi et al. 2012). Forested areas generally support higher abundances of deer ticks (Ixodes scapularis) than grasslands and shrublands but patches of little bluestem (Schizachyrium scoparium) supported small mammals with high tick burdens despite low numbers of host-seeking ticks (Ostfeld et al. 1995). It is still not clear exactly how the landscape configurations of grasslands, shrublands, and woodlands in important sandplain grassland areas influences risk of Lyme and other tick-borne diseases. One new feature of the presence of new vector-borne diseases may be the ways that landowners choose to manage their properties to reduce exposure risk. Some agencies recommend management that promotes lawns, tree removal, wood chip borders and other features to reduce tick abundance (Stafford 2002). The extent to which these land conversions from more to less natural habitat structures influence the region is not clear, but the diseases, which are the root cause of the underlying motivation to modify yards and other areas of high human contact, were not present to nearly the current extent before the significant expansion of the area of Lyme disease risk in the northeastern U.S. that occurred since Lyme disease was first described from Lyme, Connecticut in 1976.
There are other ways that managing sandplain grasslands into the future could be more difficult compared with in past decades. Vegetation management with prescribed fire today faces many challenges that include regulations under the Clean Air Act, the Endangered Species Act, other state and local regulations, and public acceptance, including perceived risks to property (Ryan et al 2013). In the densely populated northeastern U.S., and particularly in coastal locations where tourism is a major driver of local economies, burning in summer is not allowed by local fire departments. Attitudes toward prescribed fire vary widely within towns within the sandplain grassland region, adding to the challenge of applying fire when and where it would be most useful. One effect of these restrictions is that current prescribed fires, even when they can be conducted, now occur at more restricted times of the year. In the northeast, prescribed fires are conducted primarily in the spring and fall (Carlson 2013) and rarely in the summer, when the benefits to sandplain grasslands of reducing woody vegetation would be greater. Maintaining prescribed fire programs is also costly, and the unpredictability of weather, the narrowing of allowed fire windows, and greater requirements for insurance when grasslands lie adjacent to valuable coastal properties, can all add to the cost. In the future, greater precipitation—and particularly greater precipitation in late winter—may increase the difficulty of conducting prescribed fires in grasslands even during the spring and fall seasons when they typically now occur. Implementing recommendations of this review and increasing the amount of burning that occurs in sandplain grasslands during the growing season will take investments in education of both local fire departments and the public. While mowing in existing grasslands is widely accepted and faces many fewer logistical constraints, mowing does not provide some of the habitat heterogeneity and exposed soil that are benefits of prescribed burning. Mechanical clearing of forest or shrubland to expand grasslands in priority locations can also meet some public resistance. In contrast to burning, however, mechanical clearing faces fewer regulatory hurdles caused by air quality or risk to property. Objections are largely aesthetic and will likely continue to be manageable. Both prescribed fire and mowing cause some direct mortality to sandplain grassland plants and animals, including some species of conservation priority. This will continue to be a concern in the future. Applying management to only portions of conservation properties at any one time—while leaving a portion as an unburned or unmowed refuge—will remain the best strategy for managing for populations of sandplain grassland species over time.
Other Land Use Trends
New suburban residences will continue to expand onto existing unprotected open space throughout the region where sandplain grasslands occur. The rate of expansion has slowed in Massachusetts, on the Islands, and Cape Cod in recent years because of increases in land protected for conservation. The success of the local land trust movement (National Land Trust Alliance 2015) has contributed to the protection of open space that in the northeast U.S. that includes both sandplain grasslands and anthropogenic grasslands, that may have potential as sites for future sandplain grassland conversion and expansion. However, rapid expansion still continues on Long Island and in places like the interior southern counties of Massachusetts (Lautenheiser et al. 2014). Understanding and managing the multiple effects of residential development on multiple ecosystem services is a major national challenge (Groffman et al. 2017). One approach to influencing how homeowners manage residential properties is through programs such as the Habitat Network that encourages people to map and manage properties with native species to provide increased wildlife habitat and other services such as pollination. Engaging property owners around key sandplain grassland reserves could potentially reduce the harmful effects of habitat fragmentation.
There is strong interest in expanding the production of local foods across the northeast U.S. and reversing the pattern of long-term decline in the region’s farmland that has occurred since the mid-nineteenth century. The recent A New England Food Vision (Donahue et al. 2014) calls for a three-fold increase in the area of agricultural land in New England (from 2 to 6 million acres) to meet a goal of producing half of New England’s food by 2060 food while leaving 70% of the region forested. A trend in land use to accommodate greater food production could have important influences on grassland conservation and management. Because current agricultural grasslands not currently protected for conservation in the sandplain region are already cleared and because they have been in previous agricultural use, they will likely be perceived as lands onto which food production could easily be expanded. Increasing interest in use of these lands for agriculture could make it harder to expand or restore more native species-rich, sandplain grassland vegetation onto these existing grasslands in the future. If existing agricultural grasslands are maintained for grazing with a greater emphasis on food production, there is potential to make future grazing compatible with some objectives of sandplain grassland conservation, such as development of grazing systems that would promote greater cover of warm season compared with cool season grasses. Much more work needs to be conducted with local producers of grazing animals to examine the values and effects of different grazing strategies. Expanded cultivation of current agricultural grasslands would remove most of the potential for conserving the species that are sandplain grassland conservation targets in at least in the short run.
Another potential intersection of local food production and expanded local agriculture could be increased interest in using grazing animals for management of existing higher-quality conservation lands, which include currently high-quality sandplain grasslands, or sandplain grasslands into which woody vegetation is expanding. There could be both benefits and impacts of the introduction of grazing animals into conservation lands. We currently know relatively little about how grazing animals influence sandplain grassland vegetation structure, the spread of non-native species, animal habitat, or the direct disturbance effects on many important sandplain grassland species. More carefully designed and monitored experiments that examine the responses to different animals, and use of animals at different densities and different times of year, are needed.
Moving Management into the Future
The management of sandplain grasslands will have to adapt to future conditions in a number of ways. Key issues that emerge from this review that are likely to confront sandplain grassland managers in the future include the following:
(1) More aggressive approaches to controlling the regrowth of undesired woody vegetation and a wider variety of exotic and invasive species will be needed. This will require applying more treatments such as mowing, burning, soil disturbance, or herbicide applications during the growing season or applying combinations of treatments, and applying them more frequently. Many of the same approaches to woody plant management can be adapted for invasive plant management.
(2) Emphasis on conservation of larger landscapes that include sandplain grasslands as a component will be required to conserve grasslands and their associated biodiversity in the face of rapid climate change. At the scale of small regions, these landscapes should connect current grasslands near the coast with protected lands that contain grasslands, shrublands or woodlands farther inland in places that could eventually replace grasslands nearer the shore that will be lost as sea level rises and coastal flooding increase.
(3) Develop strategies for accommodating larger future variations in climate by adapting management programs, such as prescribed burning, to both allow for the flexibility of foregoing treatment in very wet years and expanding the areas treated in very dry years, when management benefits are likely to be greater. In addition, accommodation should be made to spread restoration management actions like seed collections and seeding out over different years to avoid failures during more likely but less predictable future periods of intense weather events such as rainy periods and droughts.
(4) Partnerships should be developed to test the effects of grazing animals on sandplain grasslands. This information could be used potentially to develop strategies by which grazing of animals for food production might be consistent with sandplain grassland biodiversity conservation management.
References
Aizen, M.A. & Patterson, W. A. III. 1995. Leaf phenology and herbivory along a temperature gradient: a spatial test of the phonological window hypothesis. Journal of Vegetation Science 6:543-550.
Bradley, R.S., Karmalkar, A. & K. Woods. 2018. Climate System Research Center, University of Massachsuetts, Amherst. Available at: https://www.geo.umass.edu/climate/stateclimatereports.html
Champlin, L. 2016. Quantifying rates of shrub expansion and coastal grassland loss using historical aerial imagery of Naushon Island, MA. Undergraduate thesis, Brown University. 41 pp.
Carlson, J.R. 2013. A synopsis of prescribed fire in New England. Ecological Landscape Alliance. Posted July 15, 2013. Available at: https://www.ecolandscaping.org/07/uncategorized/a-synopsis-of-prescribed-fire-in-new-england/.
DeStefano, S. & DeGraaf, R.M. 2003. Exploring the ecology of suburban wildlife. Frontiers in Ecology and Environment 1: 95-101.
Dodds, K.J., Aoki, C.F., Arango-Velez, A., Cancilliere, J. D’Amato, A.W., DeGirolomo, M.F. & Rabaglia, R.J. 2018. Expansion of southern pine beetle into Northeastern forests: Management and impact of a primary bark beetle in a new region. Journal of Forestry 116: 178-191.
Donahue, B., Burke, J., Anderson, M., Beal, A., Kell, T., Lapping, M., Rammer, H., Libby, R. & Berlin, L. 2014. A New England Food Vision. Food Solutions New England, University of New Hampshire, Durham, NH. 45 pp.
Griffiths, M. E., Keith, R. P. & Orians, C. M. 2006. Direct and indirect effects of salt spray and fire on coastal heathland plant physiology and community composition. Rhodora 108: 32-42.
Groffman, P.M., Avolio, M. Cavender-Bares, J., Bettez, N. D, Grove, J.M., Hall, S.J., Hobbie, S.E., Larson, K.L., Lerman, S.B., Locke, D.H., Heffernan, J.B., Morse, J.L., Neill, C., Nelson, K.C., O’Neil-Dunne, J., Paaki, D.E., Polsky, C., Pouyat, R., Roy Chowdhury, R., Steele, & Trammell, T.L. E. 2017. Ecological homogenization of residential macrosystems. Nature Ecology and Evolution 1, 0191. DOI:10.1038/s41559-017-0191.
Horton, R., Yohe, G., Easterling, W., Kates, R., Ruth, M., Sussman, E., Whelchel, A., Wolfe, D. & Lipschulz, F. 2014: Chapter 16: Northeast. Climate Change Impacts in the United States: The Third National Climate Assessment, Melillo, J.M., Richmond, T.C. & Yohe, G.W. (eds.), U.S. Global Change Research Program, 371-395. doi:10.7930/J0SF2T3P.
Isaksson, D., Wallander, J. & Larsson, M. 2007. Managing predation on ground-nesting birds: The effectiveness of nest exclosures. Biological Conservation 136: 136-142.
Lautzenheiser, T.E., Collins, J.M., Ricci, E.H. & Clarke, J. 2014. Losing Ground: Planning for Resilience. Massachusetts Audubon Society, Lincoln, MA. 32 pp. Available at: https://www.massaudubon.org/our-conservation-work/advocacy/shaping-the-future-of-your-community/publications-community-resources/losing-ground-report.
Levi, A., Kilpatrick, A.M., Mangel, M. &Wilmers, C.C. 2012. Deer, predators, and the emergence of Lyme disease. Proceedings of the National Academy of Sciences 109:10942-10947.
Loss, S. R., Will, T. & Mara, P.P. 2013. The impact of free-ranging domestic cats on wildlife of the United States. Nature Communications DOI:10.1038/ncomms2380.
Lutter, S. H., Dayer, A.A., Heggenstaller, E. & Larkin, J.L. 2018. Effects of biological monitoring and outreach on private landowner conservation management. PLoSONE 13: e0194740.
Miller-Rushing, A.J. & Primack, R.B. 2008. Global warming and flowering times in Thoreau’s Concord: A community perspective. Ecology 89:332-341.
Mohan, J.E., Ziska, L.H., Schlesinger, W.H., Thomas, R.B., Sicher, R.C., George K. & Clark, J.S. 2006. Biomass and toxicity responses of poison ivy (Toxicodendron radicans) to elevated atmospheric CO2. Proceedings of the National Academy of Sciences 103: 9086-9089.
Motzkin, G. & Foster, D.R. 2002. Grasslands, heathlands and shrublands in coastal New England: historical interpretations and approaches to conservation. Journal of Biogeography 29: 1569-1590.
Motzkin, G., Foster, D.R. & Ciccarello, S.C. 2002. Frost pockets on a level sandplain: Does variation in microclimate help maintain persistent vegetation pattern? Journal of the Torrey Botanical Society 129:154-163.
National Land Trust Alliance. 2015. 2015 National Land Trust Census Report. Available at: http://s3.amazonaws.com/landtrustalliance.org/2015NationalLandTrustCensusReport.pdf
Neill, C. 2017. Northward expansion of Blue Grosbeaks into Massachusetts. Bird Observer 45:377-382.
Ostfeld, R.S., Cepeda, O.M., Hazler, K.R. & Miller, M.C. 1995. Ecology of Lyme disease: Habitat associations of ticks (Ixoedes scapularis) in a rural landscape. Ecological Applications 5:353-361.
Reinmann, A.B. & Hutyra, L.R. 2017. Edge effects enhance carbon uptake and its vulnerability to climate change in temperate broadleaf forests. Proceedings of the National Academy of Sciences 114:107-112.
Ryan, K.C., Knapp, E.E. & Varner, J.M. 2013. Prescribed fire in North American forests and woodlands: history, current practice and challenges. Frontiers in Ecology and Environment 11:e15-e24.
Scaven, V. & Rafferty, N.E. 2013. Physiological effects of climate warming and insect pollinators and potential consequences for their interactions. Current Zoology 59:418-426.
Sweet, W.V., Dusek, G., Obeysekera, J. & Marra, J.J. 2018. Patterns and projections of high tide flooding along the U.S. coastline using a common impact threshold. National Oceanic and Atmospheric Administration, NOAA Technical Report NOS CO-OPS 086, Washington, D.C. 44 pp. Available at https://tidesandcurrents.noaa.gov/publications/techrpt86_PaP_of_HTFlooding.pdf
Von Holle, B. & Motzkin, G. 2007. Historical land use and environmental determinants of non-native plant distribution in coastal southern New England. Biological Conservation 136:33-43.
Vickery, P D., Hunter, M.L. & Wells, J.V. 1992. Evidence for incidental predation and its effects on nests of threatened grassland birds. Oikos 63: 281-288.
Wilson, M. L., Telford, S.R.I. III, Piesman, J. & Spielman, A. 1988. Reduced abundance of immature Ixodes dammini (Acari: Ixodidae) following elimination of deer. Journal of Medical Entomology 25:224-228.