

Conversion from Agricultural Grassland
 Figure 1. Agricultural grassland, Peterson Farm, Falmouth, MA. Photo: Chris Neill. |
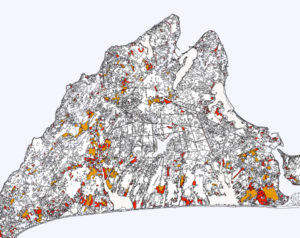 Figure 2. Area of existing sandplain grassland compared with agricultural grassland on Martha’s Vineyard. Red is high quality sandplain grassland; orange is agricultural grassland. Based on map by The Nature Conservancy (2002). |
Creating or expanding sandplain grasslands from recently-cultivated land or from agricultural grasslands that are typically pastures or hayfields (Fig. 1) is an important potential mechanism that could increase the limited area of current sandplain grasslands. Opportunities for conversion of agricultural grasslands exist because the area of these grasslands can be larger than the area of sandplain grasslands in many places (Fig. 2).
Conversion of agricultural lands or agricultural grasslands to sandplain grassland habitat aims to promote a diverse assemblage of grassland species with a high proportion of warm-season grasses and native forbs, and a low proportion of cool-season grasses and non-native invasive species. Conversion also aims to combat soil legacy effects from years of agriculture and maintain microhabitats that foster germination and regeneration of disturbance-dependent native grassland plants.
In this document, we evaluate results compiled from published and unpublished studies and information obtained from interviews with land managers. We focused on the following main questions relevant for sandplain grassland management:
1) What are the phases needed to restore sandplain grasslands on agricultural land or agricultural grasslands?
2) What management techniques have been studied?
3) What are the results of different management techniques?
4) How can the effectiveness of management techniques be improved?
We focus on interpreting the main patterns that emerge from examining multiple experiences across multiple sites, with the understanding that responses to any one treatment of a management practice under particular conditions may differ.
Methods
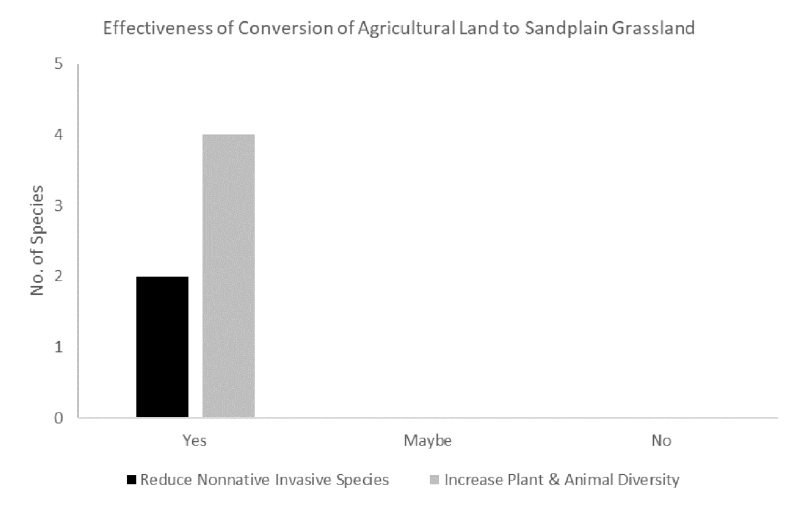
Figure 3. Number of sources that found results that suggest a decrease of non-native invasive species and/or an increase in plant and animal diversity.
We reviewed 75 sources that described or documented results of management actions in sandplain grasslands. Of these, eight sources contained information on conversion of agriculture to sandplain grasslands, and four detailed specific management experiments and case studies. In addition, we interviewed three professionals throughout the region about their experiences with conversion from agricultural land to sandplain grasslands. Literature sources that tested active management treatments were classified by whether they: (1) decreased non-native species, and (2) increased native biodiversity of plants or animals. We also used the review and interviews to summarize the state of current management practices and phases used to convert agricultural land to sandplain grasslands and their effects on: (1) fuels and soils, (2) vegetation composition, (3) vegetation structure, and (4) fauna in relation to important variables. We then suggest ways that the use of these management practices could be improved to increase warm-season graminoid cover, increase sandplain grassland forbs, and promote fauna in converted agricultural lands.
Results
Few studies have examined this conversion (Fig. 3). Well-documented regional attempts at converting recently-tilled lands or agricultural grasslands to native species-rich sandplain grasslands is limited primarily to work done on Bamford Preserve on Martha’s Vineyard and we draw heavily on those results here (Wheeler et al. 2015, Neill et al. 2015).
Conversion phases and management practices
Conversion of agricultural grasslands to sandplain grasslands typically involves three phases: (1) removal of existing non-native plants, (2) soil disturbance and/or soil amendments, and (3) seeding.
Bamford Preserve is a 25-ha property located in southeastern Martha’s Vineyard, Massachusetts owned and managed by The Nature Conservancy. The property was tilled for crops until 1992 and subsequently maintained for pasture and hay. Existing grassland was dominated by sweet vernalgrass (Anthoxanthum odoratum), orchard-grass (Dactylis glomerata), smooth brome (Bromus inermis), and non-native forbs such as ribgrass plantain (Plantago lanceolata) and Queen Anne’s lace (Daucus carota).
Existing vegetation in agricultural grasslands
Agricultural grasslands were identified as a community distinct from sandplain grasslands and heathlands by Dunwiddie et al. (1996), but can contain some plants more typical of sandplain grasslands and often include some cover of native warm-season grasses such as little bluestem (Schizachyrium scoparium), switchgrass (Panicum virgatum), or other species. However, predominant existing plant cover in recently cultivated agricultural grassland in the northeastern U.S. typically consists of non-native cool-season grasses such as A. odoratum, D. glomerata, tall rye grass (Schedonorus arundinaceus), sheep fescue (Festuca ovina), velvet-grass (Holcus lanatus), brome (Bromus spp), timothy (Phleum pretense), bentgrasses (Agrostis spp), and quack-grass (Elymus repens) (C. Neill, interview). Recently-cultivated agricultural grasslands also typically contain predominantly non-native forbs including plantains (Plantago spp.), spotted cats-ear (Hypochaeris radicata), dandelion (Taraxacum officinale), vetch (Vicia spp.), yarrow (Achillea millefolium), common sheep sorrel (Rumex acetosella), and others (C. Neill, interview). These lands are also often invaded by non-native shrubs and vines including Morrow honeysuckle (Lonicera morrowii), multiflora rose (Rosa multiflora), autumn-olive (Elaeagnus umbellata), oriental bittersweet (Celastrus orbiculatus), and increasingly in the Northeast U.S., Amur pepperbush (Ampelopsis glandulosa), and black swallow-wort (Cynanchum louiseae) (C. Neill, interview).
Phase 1: Vegetation Removal
Vegetation removal can be implemented mechanically or chemically by application of herbicide. Wheeler et al. (2015) tested five methods of removing existing vegetation at Bamford Preserve in 5 x 5-meter plots on Martha’s Vineyard: (1) tilling twice during the growing season, (2) tilling tree times during the growing season, (3) glyphosate herbicide, (4) black plastic, and (5) a Waipuna® hot foam machine designed for organic weed management of golf courses. Wheeler et al. found that tilling twice, tilling three times, black plastic and the broad-spectrum herbicide glyphosate were all roughly equally effective at reducing the formerly-established plant species and increasing the richness of native species after planting with a native seed mix. Hot foam was less effective at reducing the former species and the elimination of formerly-established plant species by herbicide resulted in slightly higher cover of native plants after five years compared with tilling and black plastic.
Phase 2: Soil Disturbance and/or Amendment

Figure 4. Soil amendment plots after tilling and treatment. Photo: Chris Neill.
Wheeler et al. (2015) also examined the effects of soil characteristics on plant recruitment and survival by testing three levels of elemental sulfur additions (to lower soil pH), three levels of wood chip additions and two levels of sawdust additions (to immobilize soil nitrogen and reduce soil available nitrogen), three levels of urea nitrogen additions (to raise soil available N to test the idea that cover of native species and native species diversity decrease with higher N), and supplemental water (to test the effect of drought stress for germination and seedling establishment) (Fig. 4).
The justifications for these treatments were: (1) lowering soil pH by adding elemental sulfur (S) has been successful in other places (Owen et al. 1999) and may improve soil conditions for restoring communities adapted to acidic soils (Walker et al. 2007); (2) the addition of organic matter with a high carbon to nitrogen ratio (C:N) has been used successfully in restoration efforts to decrease soil nitrogen because it stimulates microbial nitrogen immobilization and thereby reduces the amount of plant-available nitrogen (Morgan 1994, Blumenthal et al. 2003, Perry et al. 2010); (3) the general finding that nitrogen decreases species richness and native species abundance in many grasslands worldwide (Suding et al. 2005, Seabloom et al. 2015); and (4) the fact that even fairly dry-tolerant coastal sandplain species experienced reduced growth when water availability is very low (Griffiths and Orians 2003).
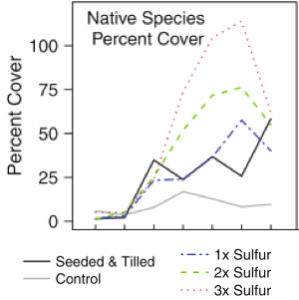
Figure 5. Response of vegetation to sulfur additions over a seven-year period. From Wheeler et al. (2015).
Wheeler et al. (2015) found that additions of carbon as sawdust or wood chips did not change soil nitrogen availability and had no effect on native cover or richness. This indicated that the amounts of these amendments that would be practical to apply as restoration management treatments had little effect on plant establishment or persistence over five years. Even high levels of nitrogen additions had little effect on soil nitrogen availability and did not affect the richness and cover of target native plant species. Additions of elemental sulfur of 91 g S/m2 reduced soil pH from 6.2 to 5.0 and a level typical of existing sandplain grassland. Sulfur additions of 182 g S/m2 and 273 g S/m2 were also applied. All three levels dramatically increased native species cover, but this effect decreased somewhat over time (Fig. 5).
Wheeler et al. (2015) found that most of this increase in sulfur caused a large increase in the cover of little bluestem (Schizachyrium scoparium) three and four years post tilling and seeding. Sulfur additions also increased the cover of hairy pinweed (Lechea mucronata), Greene’s rush (Juncus greenei), common hairgrass (Deschampsia flexuosa), big bluestem (Andropogon gerardii), switchgrass (Panicum virgatum) and frostweeds (Crocanthemum spp.). Sulfur additions reduced the abundance of ribgrass plantain (Plantago lanceolata) and smooth brome (Bromus inermis). Water additions had no effect on species richness or cover, likely because the summer in which the seeding and supplemental watering were conducted were slightly wetter than average and soil moisture was probably adequate for germination and establishment without supplemental watering.
Another important finding from Wheeler et al. (2015) was that nitrogen supply, either as increased by fertilizer additions or decreased by sawdust or wood chip additions, had no measurable effect on target species richness or cover. This indicated that nitrogen does not play a major role in structuring plant competition or causing the competitive exclusion of poorer competitors. While elevated levels of soil nitrogen are associated with declines in species richness in many grasslands (Marrs 1993, Bakker and Berendse 1999, Stevens et al. 2004, Clark and Patterson III 2007, Simkin et al. 2016), soils in coastal Northeastern grasslands may have insufficient native nitrogen for elevated nitrogen levels to cause the same kinds of effects. This would also explain why the addition of sawdust or wood chips, which have reduced non-native, invasive species abundance in other grasslands (Blumenthal et al. 2003, Burke et al. 2013), had no effect.
This conclusion in Wheeler et al. (2015) was supported by two other studies in Northeast sandplain grasslands. In one study, Weiler (2011) planted three native plant species that are targets for sandplain grassland restoration—orange milkweed (Asclepias tuberosa), little bluestem (Schizachyrium scoparium), and downy goldenrod (Solidago puberula)—into the Bamford Preserve soil treatments that included three levels of carbon addition, three levels of nitrogen addition, a control that was tilled with no amendments, and an untilled, un-manipulated control.

Figure 6. Vegetation cover monitoring at Bamford Preserve. Photo: Chris Neill.
The native species were seeded into plots that had either: (1) regenerating non-native vegetation, or (2) regenerating vegetation that was removed by clipping (as seen in Fig. 6). Clipping of surrounding vegetation increased downy goldenrod and little bluestem grass biomass within all treatments. Sulfur significantly reduced soil pH and increased the biomass of downy goldenrod and little bluestem grass. Some soil amendments increased planted native biomass somewhat compared with the control, but there were no significant differences among these treatments and the tilled control, indicating that it was tilling alone that increased native species establishment. The major conclusion of the clipping experiment was that efforts to reduce non-native species biomass will lead to more rapid establishment of native species on agricultural grasslands than will manipulation of soil properties. In a second, related study, Kinnebrew (2016) found very weak associations between soil extractable nitrogen and species richness in grassland plots on Naushon Island, MA.
Seeding
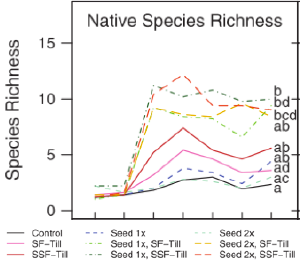
Figure 7. Tilling and seeding together increased native species richness at Bamford Preserve. From Wheeler et al. (2015).
Wheeler et al. (2015) also tested three variations of seeding with locally-collected seeds: (1) no seeding, (2) seeding in autumn in one year, and (3) seeding in autumn of two successive years. Results suggested that removal of the established plant community in combination with addition of seeds of desired species was required to increase the cover and richness of desired native sandplain grasses and forbs after five years (Fig. 7). Related experience establishing sandplain grasslands on areas cleared from forest showed that seeding led to high species richness at Job’s Neck on Martha’s Vineyard (Lezberg et al. 2006), and higher cover of native grassland species and lower cover of non-native species in coastal Connecticut (Jones et al. 2013).
A number of species appeared in main treatment plots, including in the tilled and unseeded controls that were not present in the former vegetation, the planted seed mix, or nearby surrounding lands, strongly indicating that they germinated from a soil seedbank that was established during former cultivation. The two most abundant species in this category were black mustard (Brassica nigra) and nut flatsedge (Cyperus esculentus). The presence of these plants was ephemeral and they were not present four years post-treatment.
Wheeler et al. (2015) also found that the recruitment of native species from the seedbank was not sufficient for establishing more than a small number of native species, and seeding with target plants was required. Seedbanks of native species in other agricultural grasslands are also suspected to be low although this has not been quantified. This differs from the seedbank that was found under existing species-rich sandplain grasslands on Nantucket, which were relatively rich and higher than in woodland or scrub oak shrubland (Omand et al. 2014).
Weigand (2017) studied seed limitations in the Atlantic Coastal Pine Barrens ecoregion and found that sources of seeds of desired grasses and forbs, either from a seedbank or from adjacent lands, are typically limited because of past agricultural use or habitat isolation. Thus, Weigand (2017) also argues that supplemental seeding especially of grassland specialist species is almost certainly necessary to ensure enrichment and diversification of old fields with desired species that are characteristic of regional grasslands.
The Bamford Preserve experiment used a uniform seed source to eliminate variation. However, the availability and quality of seeds, particularly local native seeds, will likely vary across years, and limit the area that can be converted and planted at one time or in one year (E. Loucks, Interview). While supplemental water did not have an effect in this experiment, drier conditions in some years may reduce germination and seedling survival.

Figure 8. Creation of warm-season grass dominated sandplain grassland at Bamford Preserve on Martha’s Vineyard. This large area was treated after evaluation of the results of multiple treatments in small plots. Photo: Chris Neill.
The findings in the small plots at Bamford Preserve were used to guide the treatment of larger areas of the existing agricultural grasslands. One subsequent attempt (Fig. 8) applied combinations of tilling and elemental sulfur additions, and was successful in 2010 (E. Loucks, Interview). Further attempts to treat larger areas in a similar manner in subsequent years (2012 and 2014) were less successful, potentially because of poor seed quality, insufficient soil-to-seed contact, seeds sinking too deep into the soil, dry conditions following seeding, or a combination of factors (E. Loucks, Interview). These attempts indicate that year-to-year variations in seed production, seed quality, and conditions during key periods of seed germination may strongly influence outcomes, even when management methods are similar.
One important unanswered question from Wheeler et al. (2015) is how the vegetation will evolve over time. There was evidence five years post-treatment that the cover of native species that had become established started to decline, and it is unknown whether these declines may influence the long-term future of plant diversity in this converted sandplain grassland. Given the history of a number of existing sandplain grasslands that occur today on formerly-tilled sites, a richer mix of native species could increase over time. This appears to have been the history at Katama Airfield and the Crane Wildlife Management Area in Falmouth, MA. The elevated pH caused by former liming declines over several decades and becomes very small after about 60 years (Neill et al. 2007), and this should favor native over non-native species. After conversion from agricultural grasslands, newly-created sandplain grasslands will require a maintenance management regime to limit regrowth of woody species and other undesirable species.
Even though the lack of a native shrub component makes sandplain grasslands created on agricultural lands different from most older, well-established grasslands, we suspect that shrub cover at Bamford Preserve will increase over time. Methods for encouraging the colonization of native shrubs but not non-native shrubs are not well known. It is not clear that acceleration even of native shrubs is desirable, given that continued expansion of shrub cover is a major management concern. Once sandplain grasslands are established, regular management of the grassland through mowing, burning, herbicide application, or other methods will almost certainly be required.
Logistical and Practical Constraints
Because agricultural lands and agricultural grasslands are already cleared and treated with conventional practices such as tilling and hay cutting, conversion of these lands to sandplain grasslands does not typically raise the same concerns as clearing of woody vegetation or disturbing former woodland to create sandplain grasslands from woodlands may. In some cases, soil disturbance can raise objections. E. Steinauer (Interview) suggests that tilling or harrowing could cause public concern in some areas on Nantucket. Use of herbicides or black plastic for vegetation removal may be controversial or undesirable in some locations. The availability of tractors and tillers to remove vegetation and prepare soils may be limited. Loss of farmland for growing local food is an increasing concern (Donahue et al. 2014).
The availability of sufficient, high-quality, native seed will likely limit future efforts to construct sandplain grasslands on formerly tilled lands or agricultural grasslands. The commercial supply of seed remains limited even with a swell of grassroot organizations and business efforts to produce local sources of genetically appropriate species for grassland restorations. Restorations now often depend on low diversity seed mixes dominated by native grass cultivars from other regions. Use of these standard seed mixes rather than local ecotypes, appears in New York to be causing the development of novel grasslands dominated by warm-season grasses and with lower grassland plant species richness (Miller 2013, Weigand 2017). Use of ecotypic, local plant materials would address this concern, but increasing the availability of local seed for sandplain grassland creation projects remains a challenge.
Another concern about the future vegetation of sandplain grasslands created on former agricultural lands is the potential invasion by a range of non-native species that are present in the landscape today but that were likely not as abundant when native grasses and forbs became established on current, but formerly-cultivated, sandplain grasslands. In addition to traditional old-field invading shrub species such as multiflora rose, Oriental bittersweet, and bush honeysuckles, new invasive shrubs and vines that are today common invaders of agricultural fields include black swallow-wort (Cynanchum louiseae), Amur pepperbush (Ampelopsis glandulosa), and spotted knapweed (Centaurea stoebe). Experience with these species in other sandplain grasslands suggests that they can be controlled in sandplain grasslands purposefully created from agricultural lands using mowing, burning, and selective removal with herbicides if a dense cover of native species is established.
Summary and Pathways to More Effective Management
This review indicated that vegetation removal and seeding are both required to establish sandplain grassland vegetation on former agricultural lands. The addition of seed alone, or removal of the formerly-established plant species alone, resulted in much lower recruitment of target plant species. Native sandplain grassland associated species increased in richness and cover after one season of tilling and seeding with native seeds. There was no clear advantage to seeding in multiple years. Tilling and seeding with native species did not eliminate formerly dominant non-native grasses and forbs, although non-native species greatly decreased in cover. The success of herbicide treatment, which did not experience soil disturbance, clearly showed that the elimination of competition with formerly-established plants, and not solely the act of physical soil disturbance by tilling, promoted germination and establishment of target plant species.
Soil amendment experiments concluded that additions of elemental sulfur of more than 91 g S/m2 reduced soil pH and dramatically increased native plant species cover, but this effect decreased somewhat over time. Additions of carbon as sawdust or wood chips also did not change soil nitrogen availability and had no effect on native cover or richness. This indicated that that amounts of these amendments that would be practical to apply as restoration management treatments had little effect on plant establishment or persistence over five years. Even high levels of nitrogen additions had little effect on soil N availability and did not affect the richness and cover of target native plant species. Lastly, nitrogen supply, either as increased by fertilizer additions or decreased by sawdust or wood chip additions, had no measurable effect on target species richness or cover. This indicated that nitrogen did not play a major role in structuring plant competition or causing the competitive exclusion of poorer competitors.
We found no research that specifically examined abundance and composition of fauna in response to creation of sandplain grasslands from agricultural lands or agricultural grasslands.
Bakker, J.P., & Berendse, F. 1999. Constraints in the restoration of ecological diversity in grassland and heathland communities. Trends in Ecology & Evolution 14: 63–68.
Blumenthal, D.M., Jordan, N.R., & Russelle, M.P. 2003. Soil carbon addition controls weeds and facilitates prairie restoration. Ecological Applications 13: 605–615.
Burke, I.C., Bontti, E.E., Barrett, J.E., Lowe, P.N., Lauenroth, W.K., & Riggle, R. 2013. Impact of labile and recalcitrant carbon treatments on available nitrogen and plant communities in a semiarid ecosystem. Ecological Applications 23: 537–545.
Clarke, G.L., & Patterson III, W.A. 2007. The distribution of disturbance-dependent rare plants in a coastal Massachusetts sandplain: Implications for conservation and management. Biological Conservation 136: 4–16.
Donahue, B., Burke, J., Anderson, M., Beal, A., Kell, T., Lapping, M., Rammer, H., Libby, R. & Berlin, L. 2014. A New England Food Vision. Food Solutions New England, University of New Hampshire, Durham, NH. 45 pp.
Dunwiddie, P.W, Zaremba, R.E., & Harper, K.A. 1996. A classification of coastal heathlands and sandplain grasslands in Massachusetts. Rhodora 98:117-145.
Griffiths, M.E., & Orians, C.M. 2003. Salt spray differentially affects water status, necrosis, and growth in coastal sandplain heathland species. American Journal of Botany 90: 1188–1196.
Jones, C.C., Dreyers, G.D., & Barrett, N. 2013. Evaluating the success of seed sowing in a New England grassland restoration. Natural Areas Journal 33: 214–221.
Kinnebrew, E. 2016. The impacts of reintroduced disturbance on soil nitrogen and vegetation composition in postagricultural grasslands. Undergraduate Thesis, University of Chicago, Chicago, IL. 57 pp.
Lezberg, A.L., Buresch, K., Neill, C., & Chase, T. 2006. Mechanical land clearing to promote establishment of coastal sandplain grassland and shrubland communities. Restoration Ecology 14: 220–232.
Marrs, R.H. 1993. Soil fertility and nature conservation in Europe: Theoretical considerations and practical management solutions (M. Begon & A. H. Fitter, Eds.). Advances in Ecological Research Volume 24: 241–300.
Morgan, J.P. 1994. Soil impoverishment a little-known technique holds potential for establishing prairie. Ecological Restoration 12:1.
Neill, C., Von Holle, B., Kleese, K., Ivy, K.D., Collins, A.R., Treat, C., & Dean, M. 2007. Historical influences on the vegetation and soils of the Martha’s Vineyard, Massachusetts coastal sandplain: Implications for conservation and restoration. Biological Conservation 136: 17–32.
Neill, C., Wheeler, M.M., Loucks, E., Weiler, A., Von Holle, B., Pelikan, M., & Chase, T. 2015. Influence of soil properties on coastal sandplain grassland establishment on former agricultural fields. Restoration Ecology 23: 531–538.
Omand, K.A., Karberg, J.M., Beattie, K.C., O’Dell, D.I., & Freeman, R.S. 2014. Soil seed bank in Nantucket’s early successional communities: implications for management. Natural Areas Journal 34: 188–199.
Owen, K.M., & Marrs, R.H. 2000. Creation of heathland on former arable land at Minsmere, Suffolk, UK: the effects of soil acidification on the establishment of Calluna and ruderal species. Biological Conservation 93: 9–18.
Perry, L.G., Blumenthal, D.M., Monaco, T.A., Paschke, M.W., & Redente, E.F. 2010. Immobilizing nitrogen to control plant invasion. Oecologia 163: 13–24.
Seabloom, E.W., Borer, E.T., Buckley, Y.M., Cleland, E.E., Davies, K.F., Firn, J., Harpole, W.S., Hautier, Y., Lind, E.M., Macdougall, A.S., Orrock, J.L., Prober, S.M., Adler, P.B., Anderson, T.M., Bakker, J.D., Biederman, L.A., Blumenthal, D.M., Stevens, C.J., Sullivan, L., Wolkovich, E., Wragg, P.D., Wright, J., & Yang, L. 2015. Plant species’ origin predicts dominance and response to nutrient enrichment and herbivores in global grasslands. Nature Communications 6: 3–8.
Simkin, S.M., Allen, E.B., Bowman, W.D., Clark, C.M., Belnap, J., Brooks, M.L., Cade, B.S., Collins, S.L., Geiser, L.H., Gilliam, F.S., Jovan, S.E., Pardo, L.H., Schulz, B.K., Stevens, C.J., Suding, K.N., Throop, H.L., & Waller, D.M. 2016. Conditional vulnerability of plant diversity to atmospheric nitrogen deposition across the United States. Proceedings of the National Academy of Sciences 113: 4086–4091.
Stevens, C.J. 2004. Impact of nitrogen deposition on the species richness of grasslands. Science 303: 1876–1879.
Suding, K.N., Collins, S.L., Gough, L., Clark, C., Cleland, E.E., Gross, K.L., Milchunas, D.G., & Pennings, S. 2005. Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proceedings of the National Academy of Sciences 102: 4387–4392.
Walker, K.J., Warman, E.A., Bhogal, A., Cross, R.B., Pywell, R.F., Meek, B.R., Chambers, B.J., & Pakeman, R. 2007. Recreation of lowland heathland on ex-arable land: Assessing the limiting processes on two sites with contrasting soil fertility and pH. Journal of Applied Ecology 44: 573–582.
Weigand, P.S., Raciti, S., Aronson, M., Senesac, M., & Krause, M. 2017. When does restoration do more harm than good? Lessons for grassland management in the Atlantic Coastal Pine Barren Ecoregion. M.S. Thesis, unpublished. Hofstra University, Long Island, N.Y.
Weiler-Lazarz, A.M. 2012. Factors limiting native species establishment on former agricultural lands. M.S. Thesis, unpublished. University of Central Florida.
Wheeler, M.M., Neill, C., Loucks, E., Weiler, A., Von Holle, B., Pelikan, M., & Chase, T. 2015. Vegetation removal and seed addition contribute to coastal sandplain grassland establishment on former agricultural fields. Restoration Ecology 23: 539–547.
Other Sources
Neill, Chris. Interview by Lena Champlin. November 2, 2016.
Loucks, Elizabeth. Interviewed by Lena Champlin. December 1, 2016.
Steinauer, Ernie. Interviewed by Lena Champlin. December 5, 2016.